| |
Результаты работы института в 2006 г.
В 2006 году Институт биофизики СО РАН выполнял НИР по следующим основным направлениям фундаментальных исследований РАН:
5.1. Структура и функции биологических макромолекул и макромолекулярных комплексов. Биокатализ.
5.10. Биохимия и физиология микрорганизмов и грибов. Использование микрогранизмов и грибов в биотехнологии.
5.11. Биохимия и физиология растений. Механизмы роста, адаптации и взаимодействия с другими организмами. Фотобиотехнология.
5.20. Организация и биосферные функции природных экосистем - лесных, морских, пресноводных и других. Динамика и механизмы устойчивости сообществ.
5.24. Воздействие факторов внешей среды на биологические системы. Радиобиология.
5.27. Оценка состояния и проблемы сохранения биоразнообразия. Мониторинг.
5.28. Научные основы рационального использования и воспроизводства биологических ресурсов.
5.30. Математические модели в биологии. Биоинформатика.
В 2006 году продолжались исследования, запланированные на период с 2002 по 2006 гг., в рамках научного направления "Биофизика и биотехнология живых систем, включая замкнутые искусственные и природные экологические системы, моделирование и прогноз их состояния" по пяти бюджетным проектам НИР Института на период 2004-2006 гг. зарегистрированым во ВНТИЦентре с присвоением шифров государственной регистрации:
Механизмы катализа в биолюминесцентных реакциях четырех типов светящихся организмов (бактерии, кишечнополостные, черви, светляки): общие закономерности и различия. Рег. номер 0120.0 404600
Исследование структурнофункциональной организации клеточной системы синтеза биополимеров полигидроксиалканоатов (ПГА) для контролируемого синтеза новых материалов и получения конструкций и препаратов медицинского назначения нового поколения. Рег. номер 0120.0 404601
Особенности миграции биологически активных веществ и элементов природного и антропогенного происхождения в трофической цепи "продуцент - консумент" в водных экосистемах бассейна Енисея (на примере жирных кислот, серы и америция-241) Рег. номер 0120.0 404602
Биотехнология рационального использования природных ресурсов: исследование структурно-функциональной организации и динамики микробных популяций, окисляющих моно- и дисульфиды восстановленных руд, содержащих благородные металлы. Рег. номер 0120.0 404603
Оценка пределов устойчивости биосферы с помощью минимальных теоретических и экспериментальных моделей, согласованных с данными глобальных наблюдений, включая спутниковые. Рег. номер 0120.0 404599
В рамках утвержденных планов НИР по проблемам биофизики экосистем и физико-химической биологии получены следующие важнейшие результаты:
По проекту "Механизмы катализа в биолюминесцентных реакциях четырех типов светящихся организмов (бактерии, кишечнополостные, черви, светляки): общие закономерности и различия". Рег. номер 0120.0 404600
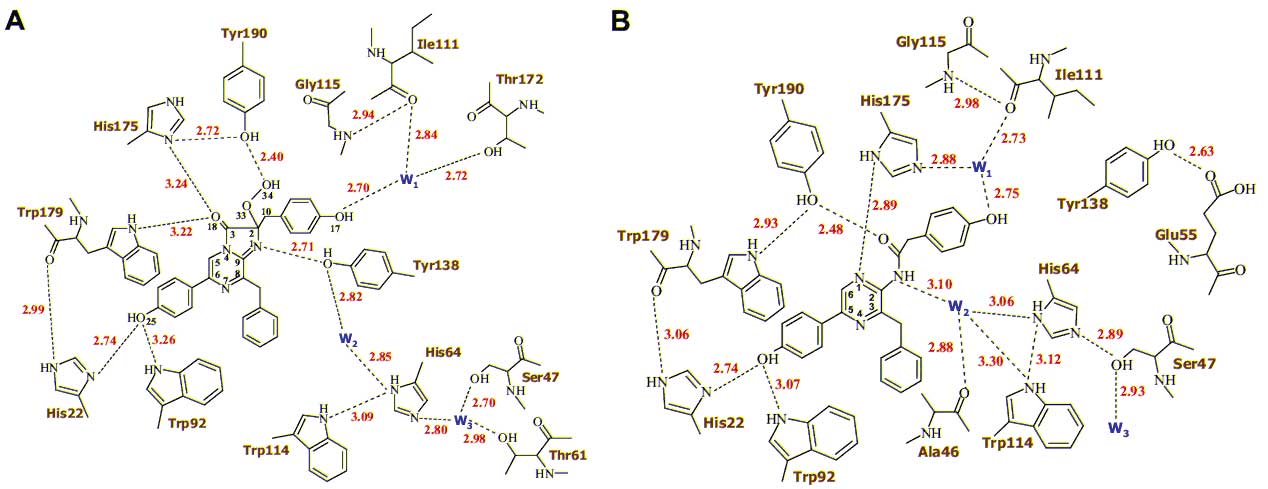
С разрешением 1.93 ? определена кристаллическая структура Са2+-разряженного фотопротеина обелина, связанного с продуктом биолюминесцентной реакции, целентерамидом, и ионами кальция. Предложен механизм образования первичного возбужденного состояния эмиттера в виде нейтральной формы целентерамида, объясняющий наблюдаемые спектры биолюминесценции.
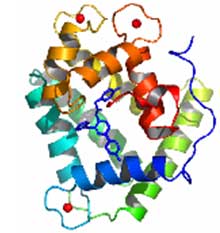
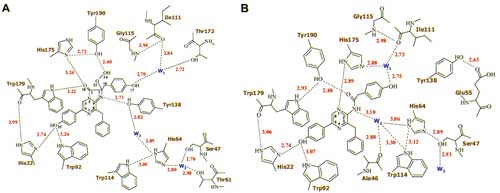
С разрешением 1.93 ? определена кристаллическая структура Са2+-разряженного фотопротеина обелина, связанного с продуктом биолюминесцентной реакции, целентерамидом, и ионами кальция (Рис. 1). Показано, что в структуре активного центра обелина (Рис. 2) лишь только два аминокислотных остатка значительно изменяют свою позицию. Имидазольное кольцо His175 становится перпендикулярным первоначальной своей ориентации, подтверждая важность данного остатка в инициации реакции, а Tyr138, ОН-группа которого формирует водородную связь с N1-атомом целентеразина в обелине, уходит из активного центра фотопротеина. На месте гидроксильной группы Tyr138 обнаружена молекула воды, которая формирует водородную связь с тем же атомом азота. На основании структуры Са2+-разряженного обелина предложен механизм образования первичного возбужденного состояния эмиттера в виде нейтральной формы целентерамида, объясняющий наблюдаемые спектры биолюминесценции.
|
Рис. 1. Структура Са2+-разряженного обелина. Молекула целентерамида показана в центре. Ионы кальция - красные шарики. Рис. 2. Распределение водородных связей (пунктирные линии) во внутренней полости обелина (A) и Ca2+-разряженного обелина (B). Видно, что после реакции водородная связь, формируемая Tyr138 с N1 атомом целентеразина (А), замещается водородной связью с молекулой воды W2 (В).
|
|
Рис. 2. Распределение водородных связей (пунктирные линии) во внутренней полости обелина (A) и Ca2+-разряженного обелина (B). Видно, что после реакции водородная связь, формируемая Tyr138 с N1 атомом целентеразина (А), замещается водородной связью с молекулой воды W2 (В).
|
При дополнительной поддержке Программы Президиума РАН и СО РАН
"Молекулярная и клеточная биология" (грант "Молекулярные механизмы
образования эмиттера в биолюминесцентных реакциях различных организмов"),
гранта РФФИ 02-04-49419. Руководитель работ: зав. лабораторией фотобиологии,
к.б.н. Е.С. Высоцкий, тел. 3912-49-44-30 Кристаллическая структура решена
совместно с Университетом штата Джорджия, США.
По проекту "Исследование структурнофункциональной организации клеточной системы синтеза биополимеров полигидроксиалканоатов (ПГА) для контролируемого синтеза новых материалов и получения конструкций и препаратов медицинского назначения нового поколения". Рег. номер 0120.0 404601
В Институте биофизики СО РАН разработана технология конструирования систем контролируемой доставки лекарственных средств с использованием полимерного резорбируемого матрикса из полигидроксиалканоатов в виде микросфер. Показана высокая противораковая эффективность разработанной экспериментальной формы высокотоксичного цитостатического препарата рубомицина на лабораторных животных с экспериментальной формой карциномы Эрлиха.
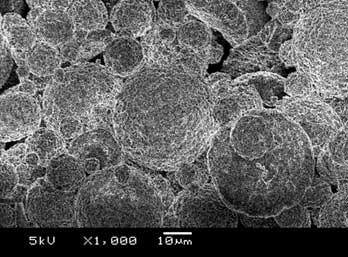
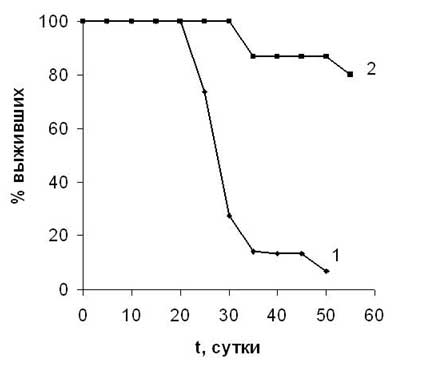
В Институте биофизики СО РАН разработана технология конструирования систем контролируемой доставки лекарственных средств (в англоязычной литературе ("drug delivery systems") с использованием полимерного резорбируемого матрикса. Получены микросферы размером от 2-3 до 25 мкм, нагруженные высокотоксичным противораковым антибиотиком антрациклинового ряда рубомицином (Рис. 3). Исследована кинетика ликвации препарата in vitro и на лабораторных животных и показано, что скорость выхода рубомицина из микросфер зависит от их структуры и нагруженности препаратом и в целом имеет длительную динамику (до 10 и более суток). На белых мышах с экспериментальной формой карциномы Эрлиха, которым внутрибрюшинно была введена суспензия раковых клеток в концентрации 5 х 103 с одновременным введением депонированного рубомицина в виде микросфер, показана высокая противораковая эффективность разработанной формы. На сроке 60 суток выживаемость животных в экспериментальной группе составила 80 % относительно 6 % - в контроле (Рис. 4).
(Гос. рег. 0120.0 404601, д.б.н. Т.Г. Волова, Лаб. хемоавтотрофного биосинтеза ИБФ СО РАН, совместно с ИФ СО РАН, Красноярским стоматологическим центром, Кардиологическим центром Красноярской краевой клинической больницы № 1, НИИ трансплантологии и искусственных органов МЗ РФ, Гематологическим научным центром РАМН).
|
Рис. 3 Микрофотографии полимерных микросфер, полученных по технологии "испарение растворителя".
|
|
Рис. 4. Динамика выживаемости лабораторных животных с экспериментальной формой
карциномы Эрлиха: 1- контрольная группа, 2-
экспериментальная группа (животные,
получившие однократно 1, 86 мг
пролонгированного рубомицина
|
(Гос. рег. 0120.0 404601, д.б.н. Т.Г. Волова, Лаб. хемоавтотрофного биосинтеза ИБФ СО РАН, совместно с ИФ СО РАН, Красноярским стоматологическим центром, Кардиологическим центром Красноярской краевой клинической больницы № 1, НИИ трансплантологии и искусственных органов МЗ РФ, Гематологическим научным центром РАМН).
По проекту "Особенности миграции биологически активных веществ и элементов природного и антропогенного происхождения в трофической цепи "продуцент - консумент" в водных экосистемах бассейна Енисея (на примере жирных кислот, серы и америция-241)" Рег. номер 0120.0 404602
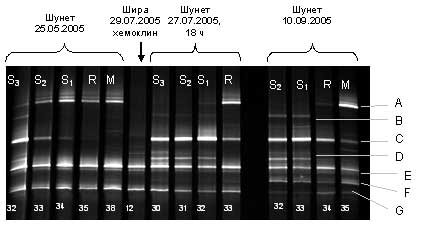
Методом ПЦР фрагментов 16S rРНК с универсальными бактериальными праймерами и последующим анализом с помощью DGGE проанализирована структура видового разнообразия и пространственного распределения доминирующих форм бактерий в зонах хемоклина озер Шира и Шунет. В хемоклине озера Шунет выявлено 7 неидентифицированных доминирующих бактериальных форм, условно обозначенных как A,B,C,D,E,F,G (Рис. 5). Обнаружено четкое пространственное разделение доминирующих форм бактерий по вертикали вдоль градиента редокс-потенциала, а также качественное изменение видового состава хемоклина оз. Шунет на протяжении исследованного периода времени - с мая по сентябрь. В бактериальном сообществе хемоклина озера Шира видны явные отличия от такового в озере Шунет: в Шира обнаружено только два доминирующих вида, E и G, причем оба этих вида входят в число доминирующих и в Шунете.
|
Рис. 5. PCR/DGGE - анализ фрагментов 16S rРНК бактериальных сообществ хемоклина озер Шира и Шунет. Обозначения дорожек вверху: М - микроаэрофильная зона (присутствуют следы кислорода), R - редокс-зона (нулевые концентрации кислорода и сероводорода), S1, S2, S3 - сероводородная зона, с увеличением номера глубина увеличивается на 5 см. Для всех серий Шунета, кроме мая, интервал глубины между соседними дорожками составляет 5 см. Для серии "Шунет 25.05.2005" различие между M и R составляет 15 см, между остальными - 5 см. Глубина во всех сериях возрастает справа налево. Дорожка для Шира получена из образца, соответствующего середине "розового" слоя со следами сероводорода. Внизу дорожек - номера шприцов пробоотборника (необходимы для точной идентификации образцов при различных анализах). (член-корр. РАН А.Г. Дегерменджи, к.ф.-м.н. Д.Ю. Рогозин)
|
По проекту "Биотехнология рационального использования природных ресурсов: исследование структурно-функциональной организации и динамики микробных популяций, окисляющих моно- и дисульфиды восстановленных руд, содержащих благородные металлы". Рег. номер 0120.0 404603
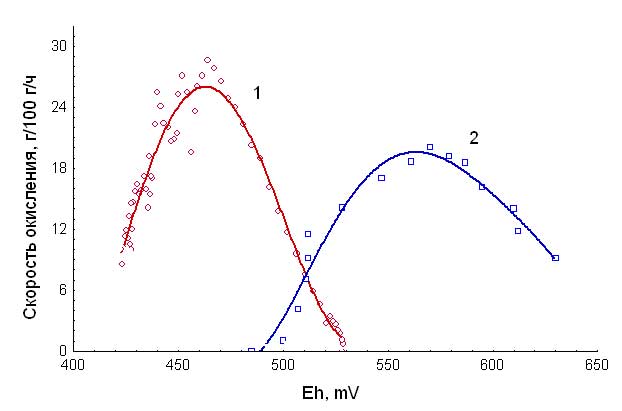
Показано, что окисление ацидофильными бактериями сульфидных минералов, которым сопутствует золото, идет в несовпадающих областях значений окислительно-восстановительного потенциала (Рис. 6). Это открывает путь к их селективному окислению и значительному уменьшению продолжительности процессов бактериальной переработки стратегического минерального сырья. (Гос. рег. 0120.0 404603, д.ф.-м.н. Ю.Л. Гуревич, лаб. Экологической биотехнологии ИБФ СО РАН).
|
Рис. 6. Зависимость скорости окисления сульфидных концентратов от окислительно-восстановительного потенциала среды (основные минералы: 1 - пирротин, 2 - арсенопирит)
|
По проекту "Оценка пределов устойчивости биосферы с помощью минимальных теоретических и экспериментальных моделей, согласованных с данными глобальных наблюдений, включая спутниковые". Рег. номер 0120.0 404599
Для увеличения дистанции и обоснованности прогноза глобальной динамики углерода требуется увеличение периода верификации минимальной модели биосферы. Увеличение дистанции прогноза требует учета вклада океана в глобальную динамику углерода, что было сделано в объединенной (суша-океан) модели. Начало периода верификации модели совпадает с началом промышленной революции 1700 года, когда было нарушено стационарное состояние углеродного цикла биосферы.
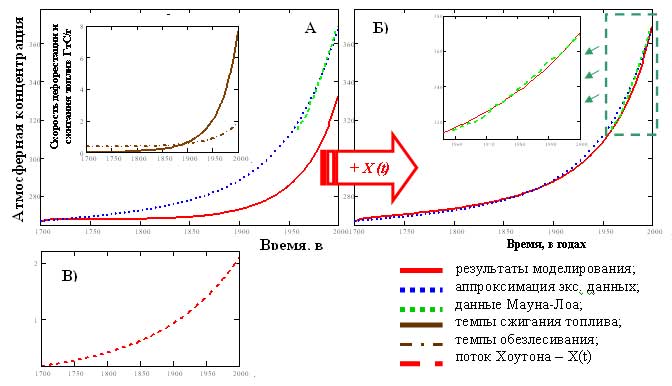
Результаты верификации объединённой многолетней модели, основанной только на данных о темпах роста сжигания топлив и обезлесивания, начиная с 1700 года, расходились с экспериментальными данными о росте концентрации углекислого газа в атмосфере (рис. 7А). Темпов сжигания ископаемых топлив оказалось недостаточно, чтобы обеспечить наблюдаемую атмосферную динамику концентрации углекислого газа. Для того чтобы привести в соответствие модельные и экспериментальные данные, требовался ещё один поток X(t) углекислого газа (рис. 7В). Величина этого потока согласуется с существующими оценками, так называемого "потока Хоутона" (~2 ГтС/г в настоящее время) (IPCC. Изменение климата, 2001; Houghton, 2002), связанного с изменением типа и способов землепользования. Введение этого потока позволило добиться хорошего соответствия модельной динамики данным наблюдения с 1700 года по наши дни (рис. 7Б).
Следует заметить, что эксперты IPCC, стараясь соблюсти баланс потоков углерода в глобальном цикле, посчитали необходимым компенсировать "поток Хоутона" неизвестным стоком в наземные экосистемы. Но как показывает данная модель, во введении неизвестного стока нет никакой необходимости.
Гос. рег. № 0120.0404599, координатор проекта член-корр. РАН А.Г.Дегерменджи, лаборатория теоретической биофизики, д.ф.- м.н. С.И.Барцев; лаборатория экологической информатики, д.т.н. А.П.Шевырногов).
|
Рис. 7. Сравнение результатов работы модели: A) модель, учитывающая только сжигание топлив и обезлесивание; Б) модель, дополнительно учитывающая неопределённый поток углерода - X(t), В) Интенсивность "потока Хоутона".
|
|